Amphibians have two types of respiratory organs (not counting the skin): gills and lungs. The weakening of gill respiration and the emergence of pulmonary respiration are already observed in Dipnoi; changes in this direction are seen in Polypterus, and Lepidosteus. In amphibians, gill respiration is preserved primarily in larvae, and then in those Urodela that spend their entire lives in water (Perennibranchiata in former systems). Gill slits are inherited by amphibians from fish-like ancestors. Gill arches are found in stegocephalians, in larvae, and in some adults (Branchiosauridae). All modern amphibians in the larval state breathe with gills. Normally, they have 5 visceral sacs and the 6th is underdeveloped. But not all of them open outward: 4 or even fewer gill slits are formed. Sometimes there are much fewer slots than arcs. The presence of cracks and arcs is proof of the origin of amphibians from fish. Internal gills, homologous to the gills of fish, are found, however, only in Anura larvae in the form of short outgrowths of integument on the arches separating the gill slits. A soft gill operculum (operculum), growing from the side of the hyoid arch, covers the gill region from the outside. The gill covers of the right and left sides merge with each other from the underside, leaving paired openings in some Anura, and one unpaired one on the left side of the body in most Anura.
In the early stages of development, the larvae of Anura and all other amphibians have only external gills, apparently homologous to the external gills of the larvae of Polypterini and Dipnoi. In Apoda and Anura, external gills exist only in the larval period, in the early stages of development, but in Urodela, which returned to aquatic life for the second time, they persist throughout life. Hence the name for these amphibians is permanent gill (Perennibranchiata), although this name, as has been said, embraces groups of amphibians having different origins. The external gills are probably inherited by amphibians from lobe-finned fishes.
Light amphibians look like long cylindrical bags with thin walls (in Urodela) or shorter ones (in Anura). In the legless, the right lung is much more developed than the left. Lungs appeared in the ancestors of tetrapods long before they landed. We see the same lungs in lungfish. They apparently appeared as an additional respiratory organ due to insufficient development of gill respiration, on the one hand, and possibly unfavorable conditions for breathing in dry and spoiled waters, on the other. The back part of the gill cavity developed in them into an additional respiratory organ. Initially, this organ, which looked like a bilobed sac that opened on the underside of the pharynx, was imperfect: its walls had to be slushy, although richly supplied with blood, with poorly developed or almost undeveloped partitions. Like all gill protrusions (slits), it had smooth visceral muscles and was innervated by the vagus first.
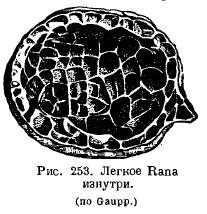
The lungs of amphibians have changed little in comparison: in aquatic Urodela, the lungs act more like a hydrostatic apparatus and have a smooth inner surface; the height of their organization is even lower than that of Dipnoi. Normally, in amphibians, the inner surface of the lungs is cellular due to the fact that a system of crossbars protrudes into the lung cavity (Fig. 253). It is very interesting that the more terrestrial a particular species is, the more developed are the crossbars in the lungs: in a toad, the lung is more cellular than frogs. In the genus Ascaphus, living in mountain streams, in oxygen-rich water, skin respiration is highly developed, while the lungs, on the contrary, are small and poorly supplied with blood. A number of amphibians from the suborder Salamandroidea (Salamandrina, Plethodon, Spelerpes, Batrachoseps, etc.) completely lost their lungs, instead of which pharyngeal and skin breathing developed strongly. .
In the simplest case, the lung sacs are interconnected in front, opening directly into the pharynx with a longitudinal slit supported on the sides by cartilaginous strips. These cartilaginous strips, with the help of muscles attached to them, can expand and narrow the laryngeal fissure.
These cartilages come from the last branchial arch and are found in their simplest form in some Urodela. From these cartilages, cartilages called cricoid cartilages can separate. They can be compared with the arytenoid cartilages (cartilagines arythenoidea) of higher vertebrates. Some Urodela and also Apoda have a rather long windpipe supported by cartilaginous rings. In Anura, the mucous membrane in the larynx forms the vocal cords. The larynx has complex muscles. At the bottom or at the corners of the mouth there are resonators that inflate when croaking.
The breathing mechanism of terrestrial amphibians is rooted in reflexes observed in fish and aquatic amphibians. Closest to fish breathing is the breathing of Anura larvae, which have internal gills, an opercular fold, and a gill cavity formed by their fusion, which opens outwards with one opening. In addition, in amphibian larvae, the oral cavity is abundantly supplied with blood. Taking water into the mouth and pushing it through the nostrils by lifting the jaws, the larvae increase the gas exchange in the oral cavity. When the larvae grow up, they rise to the surface, where they swallow air like a ceratod, and by raising the bottom of the oropharyngeal cavity they push air into the lungs. A similar act is observed in aquatic Urodela. When lowering the bottom of the oropharyngeal cavity and with the gill openings closed behind, water is sucked into the oral cavity through the mouth or nostrils, or through both. By subsequently raising the floor of the mouth with closed nostrils, water is pushed out through the gill slits. Thanks to these movements, the mucous membrane of the mouth and pharynx comes into contact with new masses of water, and the gills get a movement that renews the respiratory environment.
In terrestrial amphibians, the breathing mechanism is the act of swallowing air by lowering the muscular bottom of the mouth and pushing it into the lungs due to the bottom rising. Thus, the respiration of terrestrial amphibians is an act carried out according to the type of pressure pump that prevails in lower fish. The immediate basis on which it develops is the mechanism of respiration in perennial gill amphibians. This latter, seen in Necturus, for example, must have evolved in the distant fish-like ancestors of amphibians. A more complex type of terrestrial breathing has already developed from it - Anura.
In lungless salamanders, gas exchange of the intraoral and pharyngeal cavities is highly developed, which occurs with the help of frequent, up to 120-170 oscillations per minute of the diaphragm of the mouth (there are 30 in frogs).
In general, it should be said that lung respiration in amphibians as a whole is an auxiliary method of respiration. This also contains an indication of its phylogenetic origin.
The respiration of modern amphibians could by no means be the source of the development of respiration in the higher Tetrapoda (breathing by lifting the ribs, expanding the chest and thus sucking in air). The latter type could arise, in any case, be outlined in the most ancient extinct amphibians, which had long ribs.
Gas exchange, or respiration, is expressed in the body's absorption of oxygen from the environment (water or atmosphere) and the release of carbon dioxide into the latter as the end product of the oxidative process occurring in the tissues, due to which the energy necessary for life is released. Oxygen is taken up by the body in a variety of ways; they can basically be characterized as: 1) diffuse breathing and 2) local breathing, that is, by special organs.
diffuse breathing consists in the absorption of oxygen and the release of carbon dioxide by the entire surface of the outer cover - skin respiration - and n and e - and the epithelial membrane of the digestive tube - to and sh ch n about e respiration, i.e. without organs specially adapted for this purpose. A similar method of gas exchange is characteristic of some types of primitive multicellular animals, such as sponges, coelenterates and flatworms, and is due to their lack of a circulatory system.
It goes without saying that diffuse respiration is inherent only in organisms in which the volume of the body is small, and its surface is relatively extensive, since it is known that the volume of the body increases in proportion to the cube of the radius, and the corresponding surface - only to the square of the radius. Therefore, with a large volume of the body, this method of breathing is insufficient.
However, even with more or less appropriate volume-to-surface ratios, diffuse respiration still cannot always satisfy organisms, since the more vigorously vital activity is manifested, the more intense the oxidative processes in the body should proceed.
With intensive manifestations of life, despite the small volume of the body, it is necessary to increase its area of contact with the environment containing oxygen, and special devices to accelerate the ventilation of the respiratory tract. An increase in the area of gas exchange is achieved by the development of special respiratory organs.
Special respiratory organs vary considerably in details of construction and location in the body. For aquatic animals, such organs are the gills, for terrestrial animals, the traxae and invertebrates, and for vertebrates, the lungs.
Gill breathing. Gills are external and internal. Primitive external gills represent a simple protrusion of villous offspring of the skin, abundantly supplied with capillary vessels. Such gills in some cases differ little in their function from diffuse respiration, being only its higher stage (Fig. 332- A, 2). Usually they are concentrated in the front parts of the body.
The internal gills are formed from the folds of the mucous membrane of the initial section of the digestive tube between the gill slits (Fig. 246-2-5; 332- 7). The skin adjacent to them forms abundant branching in the form of petals with a large number of capillary blood vessels. The internal gills are often covered with a special fold of the skin (gill cover), the oscillatory movements of which improve the conditions of exchange, increasing the flow of water and removing its used portions.
Internal gills are characteristic of aquatic vertebrates, and the act of gas exchange in them is complicated by the passage of portions of water to the gill slits through the oral cavity and the movements of the gill cover. In addition, their gills are included in the circulatory circle. Each gill arch has its own vessels, and thus, at the same time, a higher differentiation of the circulatory system is carried out.
Of course, with gill methods of gas exchange, skin respiration can also be preserved, but so weak that it is relegated to the background.
In describing the oropharynx of the digestive tract, it has already been said that the gill apparatus is also characteristic of some invertebrates, such as, for example, hemichordates and chordates.
Lung breathing- a very perfect way of gas exchange, easily serving the organisms of massive animals. It is characteristic of terrestrial vertebrates: amphibians (not in the larval state), reptiles, birds and mammals. A number of organs with other functions join the act of gas exchange concentrated in the lungs, as a result of which the pulmonary method of breathing requires the development of a very complex complex of organs.
When comparing aquatic and terrestrial types of respiration in vertebrates, one important anatomical difference should be kept in mind. During gill respiration, portions of water enter the primitive mouth one by one and are released through the gill slits, where oxygen is extracted from it by the vessels of the gill folds. Thus, the gill breathing apparatus of vertebrates is characterized by an inlet and a number of outlets. During pulmonary respiration, the same openings are used for the introduction and removal of air. This feature, of course, is associated with the need to take in and push out portions of air for faster ventilation of the gas exchange area, i.e., with the need to expand and contract the lungs.
It can be assumed that the distant, more primitive ancestors of vertebrates had independent muscle tissue in the walls of the swim bladder transforming into light; with its periodic contractions, air was pushed out of the bladder, and as a result of its expansion, fresh portions of air were collected due to the elasticity of the bladder walls. Elastic tissue, along with cartilage, now dominates as a support in the respiratory system.
In the future, with an increase in the vital activity of organisms, such a mechanism of respiratory movements became already imperfect. In the history of development, it was replaced by force concentrated either in the oral cavity and the anterior part of the trachea (amphibians), or in the walls of the chest and abdominal cavities (reptiles, mammals) in the form of a specially differentiated part of the trunk muscles (respiratory muscles) and, finally, diaphragm. The lung obeys the movements of this musculature, expanding and contracting passively, and retains the elasticity necessary for this, as well as a small muscular apparatus as an auxiliary device.
 Skin respiration becomes so insignificant that its role is reduced almost to zero.
Skin respiration becomes so insignificant that its role is reduced almost to zero.
Gas exchange in the lungs in terrestrial vertebrates, as well as in aquatic ones, is closely connected with the circulatory system through the organization of a separate, respiratory, or small, circle of blood circulation.
It is quite clear that the main structural changes in the body during pulmonary respiration come down to: 1) an increase in the contact of the working area of the lungs with air, and 2) a very close and no less extensive connection of this area with the thin-walled capillaries of the circulatory circle.
The function of the respiratory apparatus - to pass air into its many channels for gas exchange - speaks for the nature of its construction in the form of an open, gaping system of tubes. Their walls, in comparison with the soft intestinal tube, are composed of a harder supporting material; in places in the form of bone tissue (nasal cavity), and mainly in the form of cartilaginous tissue and easily pliable, but quickly returning to normal elastic tissue.
The mucous membrane of the respiratory tract is lined with a special ciliated epithelium. Only in a few areas does it change into a different form in accordance with other functions of these areas, such as, for example, in the olfactory region and in the places of gas exchange itself.
Throughout the pulmonary respiratory tract, three peculiar areas attract attention. Of these, the initial - n axial strip with t - serves for the perceived air, examined here for smell. The second section - the throat - is a device for isolating the respiratory tract from the digestive tract during the passage of the food coma through the pharynx, for making sounds and, finally, for producing cough shocks that eject mucus from the respiratory tract. The last section, lёg to and e-represent the organ of direct gas exchange.
Between the nasal cavity and the larynx is the cavity of the pharynx, common with the digestive apparatus, and between the larynx and the lung, the respiratory
throat, or trachea. Thus, the passing air is used by the described expanding areas in three different directions: a) perceived odors, b) devices for making sounds and, finally, in) gas exchange, of which the latter is the main one.
The evolution of breath.
1) Diffuse breathing is the process of equalizing the concentration of oxygen inside the body and in its environment. Oxygen penetrates through the cell membrane in unicellular organisms.
2) Skin respiration- this is the exchange of gases through the skin in lower worms, in vertebrates (fish, amphibians), which have special respiratory organs.
gill breathing
PIRATE GILLS(skin outgrowths on both sides of the body) appear in marine annelids, aquatic arthropods, and mollusks in the mantle cavity.
GILLS- respiratory organs of vertebrates, formed as an invagination of the digestive tube.
In the lancelet, gill slits pierce the pharynx and open into the peribranchial cavity with frequent changes of water.
Fish have gills made from gill arches with gill filaments pierced by capillaries. The water swallowed by the fish enters the oral cavity, passes through the gill filaments to the outside, washes them and supplies the blood with oxygen.

4) Tracheal and pulmonary breathing- more efficient, since oxygen is absorbed immediately from the air, and not from the water. It is typical for terrestrial mollusks (sac-like lungs), arachnids, insects, amphibians, reptiles, birds, mammals.
arachnids have lung sacs (scorpions), tracheas (ticks), and spiders have both.

INSECTS have tracheas - the respiratory organs of terrestrial arthropods - a system of air tubes that open with breathing holes (stigmas) on the lateral surfaces of the chest and abdomen.
AMPHIBIANS have 2/3 cutaneous respiration and 1/3 pulmonary. Airways appear for the first time: larynx, trachea, bronchial rudiments; light - smooth-walled bags.
REPTILES have developed airways; the lungs are cellular, there is no skin respiration.
BIRDS have developed airways, light spongy. Part of the bronchi branches outside the lungs and forms - air sacs.

Air bags- air cavities connected to the respiratory system, 10 times the volume of the lungs, which serve to enhance air exchange in flight, do not perform the function of gas exchange. Breathing at rest is carried out by changing the volume of the chest.
Breathing in flight
1. When the wings are raised, air is sucked through the nostrils into the lungs and posterior air sacs (in the lungs I gas exchange);
Anterior air sacs ← light - posterior air sacs
2. When the wings are lowered, the air sacs are compressed, and air from the rear air sacs enters the lungs (in the lungs II gas exchange).
Front air bags - ← light rear air bags
double breath is the exchange of gases in the lungs during inhalation and exhalation.
MAMMALS- gas exchange almost entirely in the lungs (through the skin and alimentary canal -2%)

airways: nasal cavity → nasopharynx → pharynx → larynx → trachea → bronchi (bronchi branch into bronchioles, alveolar ducts and end with alveoli - pulmonary vesicles). The lungs are spongy and consist of alveoli surrounded by capillaries. The respiratory surface is increased by 50-100 times compared to the body surface. The type of breathing is alveolar. The diaphragm that separates the chest cavity from the abdominal cavity, as well as the intercostal muscles, provide ventilation to the lungs. Complete separation of the oral and nasal cavities. Mammals can breathe and chew at the same time.
Class Amphibians = Amphibians.
The first terrestrial vertebrates that still retained a connection with the aquatic environment. The class includes 3900 species and includes 3 orders: tailed (salamanders, newts), legless (tropical worms) and tailless (toads, tree frogs, frogs, etc.).
Secondary aquatic animals. Since there is no amniotic cavity in the egg (together with cyclostomes and fish, amphibians are classified as anamnia), they breed in water, where they go through the initial stages of their development. At different stages of the life cycle, amphibians lead a terrestrial or semi-aquatic lifestyle, they are distributed almost everywhere, mainly in areas with high humidity along the banks of fresh water bodies and on damp soils. Among amphibians there are no forms that could live in salty sea water. Various methods of movement are characteristic: species are known that make rather long jumps, move in steps or “crawl”, devoid of limbs (worms).
The main features of amphibians.
Amphibians have retained many features of their purely aquatic ancestors, but along with this they have also acquired a number of features characteristic of true terrestrial vertebrates.
Tailed and anurans are characterized by larval development with gill breathing in fresh water (frog tadpoles) and their metamorphosis into an adult, breathing with lungs. In legless, after hatching, the larva takes the form of an adult animal.
The circulatory system is characterized by two circles of blood circulation. The heart is three-chambered. It has one ventricle and two atria.
The cervical and sacral sections of the spine are separated, having one vertebra each.
Adult amphibians are characterized by paired limbs with articulated joints. The limbs are five-fingered.
The skull is movably articulated with the cervical vertebra by two occipital condyles.
The pelvic girdle is tightly attached to the transverse processes of the sacral vertebrae.
The eyes have movable eyelids and nictitating membranes to keep the eyes from clogging and drying out. Due to the convex cornea and flattened lens accommodation improves.
The forebrain enlarges and divides into two hemispheres. The midbrain and cerebellum are slightly developed. 10 pairs of cranial nerves leave the brain.
The skin is bare, i.e. devoid of any horny or bone formations, permeable to water and gases. Therefore, it is always moist - oxygen first dissolves in the fluid that covers the skin, and then diffuses into the blood. The same thing happens with carbon dioxide, but in reverse.
Kidneys, like in fish, primary = mesonephric.
To capture the sound waves of the air, the tympanic membrane appears, behind it is the middle ear (tympanic cavity), in which the auditory ossicle is located - the stirrup, which conducts vibrations to the inner ear. The Eustachian tube connects the middle ear cavity with the oral cavity. Choanae appear - internal nostrils, nasal passages become through.
The body temperature is unstable (poikilothermia) depends on the ambient temperature and only slightly exceeds the latter.
Aromorphoses:
Lungs and pulmonary respiration appeared.
The circulatory system has become more complex, the pulmonary circulation has developed, i.e. amphibians have two circles of blood circulation - large and small. The heart is three-chambered.
Paired five-fingered limbs were formed, which are a system of levers with articulated joints and designed for movement on land.
A cervical region was formed in the spine, which ensures the movement of the head, and a sacral region - the place of attachment of the pelvic girdle.
The middle ear, eyelids, choanae appeared.
Muscle differentiation.
Progressive development of the nervous system.
Phylogeny.
Amphibians evolved from ancient lobe-finned fish in the Devonian period of the Paleozoic era about 350 million years ago. The first amphibians - ichthyostegi - in appearance resembled modern tailed amphibians. In their structure, there were features characteristic of fish, including the rudiments of the gill cover and the organs of the lateral line.
Cover. Double layer. The epidermis is multilayered, the corium is thin, but abundantly supplied with capillaries. Amphibians retained the ability to produce mucus, but not by individual cells, as in most fish, but by formed mucous glands of the alveolar type. In addition, amphibians often have granular glands with a poisonous secret of varying degrees of toxicity. The color of the skin of amphibians depends on special cells - chromatophores. These include melanophores, lipophores and iridocytes.
Under the skin, frogs have extensive lymphatic lacunae - reservoirs filled with tissue fluid and allowing, under adverse conditions, to accumulate a supply of water.
Skeleton subdivided into axial and additional, as in all vertebrates. The vertebral column is more differentiated into sections than in fish and consists of four sections: cervical, trunk, sacral and caudal. The cervical and sacral regions each have one vertebra. Anurans usually have seven trunk vertebrae, and all the caudal vertebrae (about 12) merge into a single bone - the urostyle. Caudates have 13 - 62 trunk and 22 - 36 tail vertebrae; in legless, the total number of vertebrae reaches 200 - 300. The presence of a cervical vertebra is important, because. unlike fish, amphibians cannot turn their body so quickly, and the cervical vertebra makes the head mobile, but with a small amplitude. Amphibians cannot turn their heads, but they can tilt them.
The vertebrae of different amphibians can differ in their type. In legless and lower caudate vertebrae, the vertebrae are amphicoelous, with a preserved notochord, like in fish. The higher caudate vertebrae are opisthocoelous, i.e. the bodies are concave in front and concave in the back. In anurans, on the contrary, the anterior surface of the vertebral bodies is concave, and the posterior surface is convex. Such vertebrae are called procoelous. The presence of articular surfaces and articular processes provides not only a strong connection of the vertebrae, but also makes the axial skeleton mobile, which is important for the movement of caudal amphibians in water without the participation of limbs, due to the lateral bends of the body. In addition, vertical movements are possible.
The amphibian skull is like a modified skull of a bony fish, adapted to terrestrial existence. The brain skull remains predominantly cartilaginous for life. The occipital region of the skull contains only two lateral occipital bones, which are carried along the articular condyle, with the help of which the skull is attached to the vertebrae. The visceral skull of amphibians undergoes the greatest transformations: secondary upper jaws appear; formed by the intermaxillary (premaxillary) and maxillary bones. The reduction of gill breathing led to a radical change in the hyoid arch. The hyoid arch is transformed into an element of the hearing aid and a hyoid plate. Unlike fish, the visceral skull of amphibians is directly attached by palatal-square cartilage to the bottom of the brain skull. This type of direct connection of the components of the skull without the participation of elements of the hyoid arch is called autostyle. Gill cover elements are absent in amphibians.
The accessory skeleton includes the bones of the girdles and free limbs. As in fish, the bones of the shoulder girdle of amphibians are located in the thickness of the muscles that connect them to the axial skeleton, but the girdle itself is not directly connected to the axial skeleton. The belt provides support for the free limb.
All land animals constantly have to overcome gravity, which fish do not need to do. The free limb serves as a support, allows you to raise the body above the surface and provides movement. Free limbs consist of three sections: proximal (one bone), intermediate (two bones) and distal (relatively large number of bones). Representatives of different classes of terrestrial vertebrates have structural features of one or another free limb, but all of them are of a secondary nature.
In all amphibians, the proximal section of the free forelimb is represented by the humerus, the intermediate section by the ulna and radius in caudates, and a single forearm bone (it is formed as a result of fusion of the ulna and radius) in anurans. The distal section is formed by the carpus, metacarpus and phalanges of the fingers.
The belt of the hind limbs articulates directly with the axial skeleton, with its sacral section. A reliable and rigid connection of the pelvic girdle with the spinal column provides the work of the hind limbs, which are more important for the movement of amphibians.
Muscular system different from the muscular system of fish. Trunk musculature retains a metameric structure only in legless ones. In caudate amphibians, the metamerism of segments is disturbed, while in anurans, segments of muscle segments begin to separate, differentiating into ribbon-like muscles. The mass of muscles of extremities sharply increases. In fish, the movements of the fins are provided mainly by the muscles located on the body, while the five-fingered limb moves due to the muscles located in itself. A complex system of muscles appears - antagonists - flexor and extensor muscles. Segmented muscles are present only in the region of the spinal column. The muscles of the oral cavity (chewing, tongue, floor of the mouth) become more complex and specialized, not only participating in the capture and swallowing of food, but also providing ventilation of the oral cavity and lungs.
body cavity- in general. In amphibians, the relative position of the pericardial cavity has changed due to the disappearance of the gills. She was pushed to the bottom of the chest in the area covered by the sternum (or coracoid). Above it, in a pair of coelomic canals, lie the lungs. Cavities containing the heart and lungs. Separated by the pleurocardial membrane. The cavity in which the lungs are located communicates with the main coelom.
Nervous system. The brain is of the ichthyopsid type, i.e. the main integrating center is the midbrain, but the amphibian brain has a number of progressive changes. The amphibian brain has five sections and differs from the fish brain mainly in the large development of the forebrain, the complete separation of its hemispheres. In addition, the nerve substance already lines, in addition to the bottom of the lateral ventricles, the sides and the roof, forming the cerebral vault - the archipallium. The development of the archipallium, accompanied by increased connections with the diencephalon and especially the midbrain, leads to the fact that the associative activity that regulates behavior in amphibians is carried out not only by the medulla oblongata and midbrain, but also by the hemispheres of the forebrain. The elongated hemispheres in front have a common olfactory lobe, from which two olfactory nerves originate. Behind the forebrain is the diencephalon. On its roof is the epiphysis. On the underside of the brain is the optic chiasm (chiasm). The funnel and pituitary gland (lower brain gland) depart from the bottom of the diencephalon.
The midbrain is represented by two round optic lobes. Behind the visual lobes lies an underdeveloped cerebellum. Immediately behind it is the medulla oblongata with a rhomboid fossa (fourth ventricle). The medulla oblongata gradually passes into the spinal cord.
In amphibians, 10 pairs of head nerves leave the brain. The eleventh pair is not developed, and the twelfth departs outside the skull.
There are 10 pairs of true spinal nerves in a frog. The anterior three take part in the formation of the brachial plexus, which innervates the forelimbs, and the four posterior pairs take part in the formation of the lumbosacral plexus, which innervates the hind limbs.
sense organs provide orientation of amphibians in water and on land.
Lateral line organs are present in all larvae and in adults with an aquatic lifestyle. Represented by a cluster of sensitive cells with nerves suitable for them, which are scattered throughout the body. Sensitive cells perceive temperature, pain, tactile sensations, as well as changes in humidity and chemical composition of the environment.
Organs of smell. Amphibians have a small external nostril on each side of the head, which leads to an elongated sac ending in an internal nostril (choana). Choanae open in front of the roof of the oral cavity. In front of the choanae, on the left and on the right, there is a bag that opens into the nasal cavity. This is the so-called. vomeronasal organ. It contains a large number of sensory cells. Its function is to receive olfactory information about food.
The organs of vision have a structure characteristic of a terrestrial vertebrate. This is expressed in the convex shape of the cornea, the lens in the form of a biconvex lens, in the movable eyelids that protect the eyes from drying out. But accommodation, as in fish, is achieved by moving the lens by contracting the ciliary muscle. The muscle is located in the annular ridge surrounding the lens, and when it contracts, the frog lens moves forward somewhat.
The organ of hearing is arranged according to the terrestrial type. The second section appears - the middle ear, in which the auditory ossicle, the stirrup, which first appears in vertebrates, is placed. The tympanic cavity is connected to the pharyngeal region by the Eustachian tube.
The behavior of amphibians is very primitive, conditioned reflexes are developed slowly, and fade quickly. The motor specialization of reflexes is very small, so the frog cannot form a protective reflex of withdrawal of one leg, and when one limb is stimulated, it pulls with both legs.
Digestive system begins with the oral fissure leading to the oropharyngeal cavity. It has a muscular tongue. The ducts of the salivary glands open into it. The tongue and salivary glands first appear in amphibians. The glands serve only to moisten the food bolus and are not involved in the chemical processing of food. On the premaxillary, maxillary bones, the vomer, there are simple conical teeth, which are attached to the bone with their base. The digestive tube is differentiated into the oropharyngeal cavity, a short esophagus, which performs the function of carrying food into the stomach, and a voluminous stomach. Its pyloric part passes into the duodenum - the beginning of the small intestine. The pancreas lies in the loop between the stomach and the duodenum. The small intestine smoothly passes into the large intestine, which ends with a pronounced rectum, which opens into the cloaca.
The digestive glands are the liver with the gallbladder and the pancreas. The ducts of the liver, together with the duct of the gallbladder, open into the duodenum. The ducts of the pancreas flow into the duct of the gallbladder, i.e. This gland has no independent communication with the intestines.
That. The digestive system of amphibians differs from the similar system of fish in the greater length of the digestive tract, the final section of the large intestine opens into the cloaca.
Circulatory system closed. Two circles of blood circulation. The heart is three-chambered. In addition, there is a venous sinus in the heart, which communicates with the right atrium, and an arterial cone departs from the right side of the ventricle. Three pairs of vessels depart from it, homologous to the gill arteries of fish. Each vessel begins with an independent opening. All three vessels of the left and right sides go first as a common arterial trunk, surrounded by a common membrane, and then branch out.
The vessels of the first pair (counting from the head), homologous to the vessels of the first pair of branchial arteries of fish, are called carotid arteries, which carry blood to the head. Through the vessels of the second pair (homologous to the second pair of gill arteries of fish) - the aortic arches - the blood is directed to the back of the body. Subclavian arteries depart from the aortic arches, carrying blood to the forelimbs.
Through the vessels of the third pair, homologous to the fourth pair of gill arteries of fish - the pulmonary arteries - blood is sent to the lungs. A large cutaneous artery departs from each pulmonary artery and sends blood to the skin for oxidation.
Venous blood from the anterior end of the body is collected through two pairs of jugular veins. The latter, merging with the skin veins, which have already taken in the subclavian veins, forms two anterior vena cava. They carry mixed blood into the venous sinus, since arterial blood moves through the skin veins.
Amphibian larvae have one circle of blood circulation, their circulatory system is similar to the circulatory system of fish.
Amphibians have a new circulatory organ - the red bone marrow of tubular bones. Erythrocytes are large, nuclear, leukocytes are not the same in appearance. There are lymphocytes.
Lymphatic system. In addition to the lymphatic sacs located under the skin, there are lymphatic vessels and hearts. One pair of lymphatic hearts is placed near the third vertebra, the other - near the cloacal opening. The spleen, which looks like a small round body of red color, is located on the peritoneum near the beginning of the rectum.
Respiratory system. Fundamentally different from the respiratory system of fish. In adults, the respiratory organs are the lungs and skin. The respiratory tract due to the absence of the cervical region is short. Represented by the nasal and oropharyngeal cavities, as well as the larynx. The larynx opens directly into the lungs with two openings. Due to the reduction of the ribs, the lungs are filled by swallowing air - according to the principle of a pressure pump.
Anatomically, the amphibian respiratory system includes the oropharyngeal cavity (upper airways) and the larynx-tracheal cavity (lower tracts), which directly passes into the saccular lungs. The lung in the process of embryonic development is formed as a blind outgrowth of the anterior (pharyngeal) section of the digestive tube, therefore, in the adult state, it remains connected with the pharynx.
That. The respiratory system in terrestrial vertebrates is anatomically and functionally divided into two sections - the airway system and the respiratory section. The airways carry out two-way air transport, but do not participate in gas exchange itself, the respiratory section carries out gas exchange between the internal environment of the body (blood) and atmospheric air. Gas exchange occurs through the surface liquid and proceeds passively in accordance with the concentration gradient.
The system of gill covers becomes unnecessary, so the gill apparatus in all terrestrial animals is partially modified, its skeletal structures are partly part of the skeleton (cartilage) of the larynx. Ventilation of the lungs is carried out due to the forced movements of special somatic muscles in the process of the respiratory act.
excretory system, as in fish, it is represented by primary, or trunk kidneys. These are compact, reddish-brown bodies, lying on the sides of the spine, and not ribbon-like, like in fish. From each kidney, a thin Wolf canal extends to the cloaca. In female frogs, it serves only as a ureter, while in males it serves as both an ureter and a vas deferens. In the cloaca, the Wolf channels open with independent openings. It also opens separately into the cloaca and bladder. The final product of nitrogen metabolism in amphibians is urea. In aquatic amphibian larvae, the main product of nitrogen metabolism is ammonia, which is excreted in the form of a solution through the gills and skin.
Amphibians are hyperosmotic animals with respect to fresh water. As a result, water constantly enters the body through the skin, which does not have mechanisms to prevent this, as in other terrestrial vertebrates. Sea water is hyperosmotic in relation to the osmotic pressure in the tissues of amphibians; when they are placed in such an environment, water will leave the body through the skin. That is why amphibians cannot live in sea water and die in it from dehydration.
Sexual system. In males, the reproductive organs are represented by a pair of rounded whitish testes adjacent to the ventral surface of the kidneys. From the testes to the kidneys stretch thin vas deferens. Sexual products from the testis through these tubules are sent to the bodies of the kidneys, then to the Wolf channels and through them to the cloaca. Before flowing into the cloaca, the Wolfian channels form a small expansion - the seminal vesicles, which serve for the temporary deposition of sperm.
The reproductive organs of females are represented by paired ovaries with a granular structure. Above them are fat bodies. They accumulate nutrients that ensure the formation of reproductive products during hibernation. In the lateral parts of the body cavity there are strongly convoluted light oviducts, or Müllerian canals. Each oviduct into the body cavity in the region of the heart opens with a funnel; the lower uterine part of the oviducts is sharply expanded and opens into the cloaca. Ripe eggs through the rupture of the walls of the ovary fall into the body cavity, then are captured by the funnels of the oviducts and move along them to the cloaca.
Wolf channels in females perform only the functions of the ureters.
In tailless amphibians, fertilization is external. The eggs are immediately irrigated with seminal fluid.
External sexual characteristics of males:
Males have a genital wart on the inner finger of the forelimbs, which reaches a special development by the time of reproduction and helps the males to hold the females during the fertilization of eggs.
Males are usually smaller than females.
Development amphibians is accompanied by metamorphosis. Eggs contain relatively little yolk (mesolecithal eggs), so radial cleavage occurs. A larva emerges from the egg - a tadpole, which in its organization is much closer to fish than to adult amphibians. It has a characteristic fish-like shape - a long tail surrounded by a well-developed swimming membrane, on the sides of the head it has two or three pairs of external feathery gills, paired limbs are absent; there are organs of the lateral line, the functioning kidney is the pronephros (pronephros). Soon the external gills disappear, and in their place three pairs of gill slits with their gill filaments develop. At this time, the similarity of the tadpole with the fish is also a two-chambered heart, one circle of blood circulation. Then, by protrusion from the abdominal wall of the esophagus, paired lungs develop. At this stage of development, the arterial system of the tadpole is extremely similar to the arterial system of the lobe-finned and lungfish, and the only difference is that, due to the absence of the fourth gill, the fourth afferent branchial artery passes into the pulmonary artery without interruption. Even later, the gills are reduced. A fold of skin is formed in front of the gill slits on each side, which, gradually growing back, tightens these slits. The tadpole switches entirely to pulmonary breathing and swallows air through its mouth. Subsequently, paired limbs are formed in the tadpole - first the front, then the hind. However, the anterior ones are hidden under the skin longer. The tail and intestines begin to shorten, mesonephros appears, the larva gradually passes from plant food to animal food and turns into a young frog.
During the development of the larva, its internal systems are rebuilt: respiratory, circulatory, excretory, digestive. Metamorphosis ends with the formation of a miniature copy of an adult.
Ambistomas are characterized by neoteny, i.e. they breed larvae, which for a long time were taken for an independent species, so they have their own name - axolotl. Such a larva is larger than an adult. Another interesting group is the water-dwelling proteas, which retain their external gills throughout their lives, i.e. signs of a larva.
The metamorphosis of a tadpole into a frog is of great theoretical interest, since not only proves that amphibians evolved from fish-like creatures, but makes it possible to restore in detail the evolution of individual organ systems, in particular the circulatory and respiratory systems, during the transition of aquatic animals to terrestrial ones.
Meaning amphibians is that they eat many harmful invertebrates and themselves serve as food for other organisms in food chains.
Evolution of the respiratory system
Stages of the breathing process
Breath- a set of processes that ensure the supply of oxygen from the environment to the body, which is necessary for the oxidation of organic substances in the mitochondria of the cell, and the release of carbon dioxide
Breath types:

Breath type:
Cellular.
Organisms: unicellular animals (amoeba, green euglena, infusoria slipper); coelenterates (jellyfish, coral polyps); some worms.
Single-celled organisms absorb oxygen dissolved in water over the entire surface of the body by diffusion.
Oxygen is involved in the breakdown of complex organic substances, resulting in the release of energy, which is necessary for the life of the animal.
Carbon dioxide formed as a result of respiration is also released outward through the entire surface of the body.
Tracheal breathing is breathing with the help of a system of combined tracheal tubes that permeate the entire body.
Organisms: class Insects (beetles, butterflies, grasshoppers, flies)
The abdomen of an insect is divided into 5–11 parts (segments). Each of them has a pair of small holes - spiracle. From each spiracle branching tubules extend inward - trachea that permeate the entire body of the insect. Watching the cockchafer, you can see how its abdomen either decreases in volume or increases. These are breathing movements. When you inhale, air containing oxygen enters the body through the spiracles, and when you exhale, air saturated with carbon dioxide leaves.
In spiders (class Arachnids), the respiratory organs are represented not only by tracheae, but also by lung sacs that communicate with the external environment through respiratory openings.
Gill breathing is breathing with the help of specialized formations with a dense network of blood vessels.
Organisms: many aquatic life (fish, crayfish, mollusks)
Fish breathe oxygen dissolved in water with the help of special branched skin outgrowths called gills.
Fish are constantly swallowing water. From the oral cavity, water passes through the gill slits, wash the gills and exits from under the gill covers. Gills consist of gill arches and gill filaments which are pierced by many blood vessels. From the water that washes the gills, oxygen enters the blood, and carbon dioxide is removed from the blood into the water. The gills inside the body are called internal gills.
Some animals, such as amphibians, have thick tufts of gills on the surface of the body. Such gills are called - outdoor. Such is the structure of Proteus, a blind cave animal from the western regions of Yugoslavia, and axolotls (which are similar in general appearance to newts) - their homeland is Mexico.